Antatt fusjonssted inneholder uttrykte gener, påvirker sannsynligvis nevral utvikling
 Av Casey Luskin 16. februar 2026. Oversatt herfra. {kursiv og understreking ved oversetter.}
Av Casey Luskin 16. februar 2026. Oversatt herfra. {kursiv og understreking ved oversetter.}
Menneskelig kromosomfusjon har ofte blitt sitert som sterke bevis som støtter felles avstamning mellom menneske og ape. Det er det ikke. Som en fersk artikkel i Cell Genomics rapporterer: "Ufullstendig avstamningssortering av segmentale duplikasjoner definerer fusjonsstedet for menneskelig kromosom 2 tidlig under artsdannelsen i Afrika." Den gir noen nye avsløringer om det antatte fusjonsstedet i menneskelig kromosom 2.
Vi har skrevet om dette emnet tidligere (se for eksempel her, her og her). Som jeg har sagt før, selv om vi fullt ut innrømmer at menneskelig kromosom 2 faktisk er et resultat av fusjonen av to kromosomer, viser det bare at et sted innenfor vår menneskelige avstamning skjedde en kromosomfusjonshendelse. Om avstamningen vår kan spores tilbake til en felles avstamning med aper er et helt annet spørsmål, som ikke avgjøres av kromosomfusjon.
Bilde 1. Sjimpansen har 48 kromosomer (24 par)
Artikkel gir en barokk forklaring
Nå har jeg aldri følt sterke motiver for å tvile på menneskelig kromosomfusjon - hvis det viser seg å være sant, så er jeg glad for å akseptere det. Likevel har det blitt reist noen merkelige bevis som setter det i tvil, i hvert fall til en viss grad. Fan et al. (2002) fant at det antatte fusjonsstedet er "betydelig degenerert" og "svært avvikende fra de prototypiske telomeriske repetisjonene" sammenlignet med hva man ville forvente av et perfekt fusjonssted. Denne nye artikkelen i Cell Genomics prøver å forklare hvorfor "fusjonsstedet" er så forvrengt, og forteller en svært innviklet mutasjonshistorie for å forklare dets nåværende "degenererte" tilstand. Her er den korte versjonen av det:
"Fusjonen var assosiert med flere perisentriske inversjoner, segmentale duplikasjoner (SD-er) og omsetning av subterminalt repeterende DNA."
 Dette virker i samsvar med det vi har hørt før om fusjonsstedet. Jeg skrev om Fan et al. (2002) for år siden her og bemerket at artikkelen til og med stilte spørsmålet: "Hvis fusjonen skjedde innenfor de telomeriske repetisjonsmatrisene for mindre enn ~6 mya [millioner år siden], hvorfor er matrisene på fusjonsstedet så degenererte?" Det er fortsatt et godt spørsmål - svarer denne nye artikkelen på det?
Dette virker i samsvar med det vi har hørt før om fusjonsstedet. Jeg skrev om Fan et al. (2002) for år siden her og bemerket at artikkelen til og med stilte spørsmålet: "Hvis fusjonen skjedde innenfor de telomeriske repetisjonsmatrisene for mindre enn ~6 mya [millioner år siden], hvorfor er matrisene på fusjonsstedet så degenererte?" Det er fortsatt et godt spørsmål - svarer denne nye artikkelen på det?
Ikke på noen elegant måte. Historien som fortelles i den nye artikkelen er fryktelig komplisert og ugjennomsiktig. Jeg er ikke sikker på om den er noe mer enn bare en gigantisk, barokk, en i ettertid 100 prosent evolusjonsbasert mutasjonshistorie, som 'bare er slik'. Denne typen historier er vanskelige å tilbakevise, men får en også til å lure på om de virkelig er det riktige svaret, eller om noe annet kan være på ferde. Bortsett fra alt dette, tilbyr artikkelen noe fascinerende ny informasjon om det antatte fusjonsstedet: det inneholder uttrykte gener som ser ut til å være funksjonelle, og ser ut til å være assosiert med viktige menneskelige nevrale funksjoner.
Bilde 2. Kromosomtall hos ulike familier
Funksjonelle gener på fusjonsstedet
Artikkelen rapporterer at fusjonsstedet kan være funksjonelt, og forfatterne støtter denne påstanden med funnet at det inneholder fire gener/pseudogener som uttrykkes i forskjellige menneskelige vev:
Siden fusjonsstedet er nesten fiksert i det menneskelige genomet, spekulerte vi i at det kan gi en funksjonell fordel. Fire antatte ikke-kodende gener/pseudogener (PGM5P4, FAM138B, WASH2P og DDX11L2) har blitt annotert på stedet for den menneskelige chr2-fusjonen. I følge GTEx short-read RNA-sekvensering (RNA-seq) uttrykkes disse fire genene/pseudogenene i testikel-, spiserør-, eggleder- og lillehjernevev. Videre støttes både PGM5P4 og WASH2P av transkriptdata for long-read isoformsekvensering (Iso-Seq) fra CHM13hTERT og nyrevev fra ENCODE. I tillegg avgrenser metyleringsanalyse en fremtredende CpG-øy som viser promotorere/forsterkere av PGM5P4, identifisert ved hjelp av ONT-avlesninger fra T2T-CHM13-cellelinjen.
Ikke gå glipp av frasen om at fusjonsstedet "kan gi en funksjonell fordel" - forfatterne tar dets universelle persistens i det menneskelige genomet som bevis på at det er der fordi det gjør noe nyttig!
 Potensiell involvering i nevronutvikling
Potensiell involvering i nevronutvikling
Enda mer merkelig er det at forskerne deretter slo ut fusjonsstedet i visse celler og fant endringer i "transkripsjonsprogrammer involvert i nevronutvikling":
Differensiell genuttrykksanalyse identifiserte 547 oppregulerte og 869 nedregulerte gener ved hjelp av standardinnstillinger i den differensielt uttrykte genpipelinen (DEG). .. Funksjonell berikelsesanalyse viste at de oppregulerte genene var signifikant assosiert med mønsterspesifikasjonsprosesser (justert p = 8,61  10^5), mens de nedregulerte genene var beriket for veier relatert til nevral utvikling og organisering, inkludert forhjerneutvikling (justert p = 8,02 10^15) og aksonogenese (justert p = 4,92 10^14). Disse funnene tyder på at sletting av fusjonsstedet kan påvirke transkripsjonsprogrammer involvert i nevral utvikling, og potensielt bidra til fenotypisk divergens mellom mennesker og ikke-menneskelige primater.
10^5), mens de nedregulerte genene var beriket for veier relatert til nevral utvikling og organisering, inkludert forhjerneutvikling (justert p = 8,02 10^15) og aksonogenese (justert p = 4,92 10^14). Disse funnene tyder på at sletting av fusjonsstedet kan påvirke transkripsjonsprogrammer involvert i nevral utvikling, og potensielt bidra til fenotypisk divergens mellom mennesker og ikke-menneskelige primater.
Bilde 3. Nevroner danner ofte nett
Hvis dette fusjonsstedet har store effekter på transkripsjon relatert til nevral utvikling, høres det definitivt ikke ut som en tilfeldig søppelregion som følge av den vilkårlige fusjonen av to kromosomer som deretter opplevde alle slags komplekse mutasjoner de siste millionene av årene. Det virker som noe langt viktigere - noe som til og med kan vise tegn på design.
Fylogenetiske konflikter
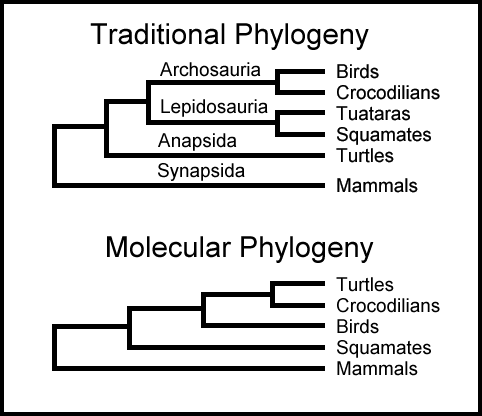 Et annet interessant funn fra denne artikkelen er at når det antatte fusjonsstedet sammenlignes med tilsvarende genomiske seksjoner hos aper, finnes det likhetsmønstre som motsier standardfylogenien til store aper og mennesker. De fant én region i det menneskelige fusjonsstedet som ligner mer på gorilla enn på sjimpanse eller bonobo:
Et annet interessant funn fra denne artikkelen er at når det antatte fusjonsstedet sammenlignes med tilsvarende genomiske seksjoner hos aper, finnes det likhetsmønstre som motsier standardfylogenien til store aper og mennesker. De fant én region i det menneskelige fusjonsstedet som ligner mer på gorilla enn på sjimpanse eller bonobo:
-Det er verdt å merke seg at SD_fusion_A (chr2) og andre menneskelige kopier deler en monofyletisk opprinnelse (3,3 millioner år), nærmest beslektet med gorillakopier ca. 6 millioner år siden (95 % konfidensintervall [KI]: 4,8-7,4 millioner år siden) i stedet for sjimpanse eller bonobo - et mønster som er i samsvar med Ufullstendig AvstamningSsortering (UAS).
Bilde 4. Fylogeni-resultater avhenger av metode
Standard fylogeni hevder selvfølgelig at mennesker er nærmere beslektet med sjimpanser enn med gorillaer, så dette passer ikke med det forventede fylogenetiske treet. De påberoper seg Ufullstendig AvstamningSsortering (UAS) for å forklare denne avviken, men det er bare en episyklus som vi ofte har sett påberopt av evolusjonsbiologer når noen genetiske data ikke passer med det forventede fylogenetiske treet (se for eksempel her). De tilskriver disse særegne fylogenetiske dataene til UAS, men UAS er i bunn og grunn en forklaring som brukes for å bortforklare fylogenetisk inkongruens.
Deretter påberoper de seg en annen forekomst av UAS (som egentlig bare betyr "fylogenetisk inkongruens") for å forklare hvorfor en annen del av genomet er mer likt gorilla hos sjimpanse/bonobo enn hos menneske:
"På samme måte er SD_fusion_B et 36 kbp-segment som tilsvarer det delvis avkortede WASH245 (11 eksoner av et forfedresgen som potensielt regulerer aktincytoskjelett-dynamikk), FAM138B (3 eksoner av en (forfedregen med ukjent funksjon), og DDX11 (3 eksoner av et forfedresgen involvert i DNA-metabolisme). Segmentet finnes som en enkelt kopi i både makaker og orangutanger og stammer fra en lokus-kartlegging til forfedres afrikansk storape chr12 (fylogenetisk gruppe XII). Alle menneskelige kopier som kartlegges til menneskelig chr2, chr20, chr12 og chr9 viser en monofyletisk opprinnelse (ca.4,4 millioner år siden, 95 % KI (KonfidensInterval): 3,1-5,8 millioner år siden), noe som tyder på menneske-spesifikke duplikasjoner eller interlokus-genkonverterings-hendelser. Med hensyn til ikke-menneskelige afrikanske storeaper avviker imidlertid denne kladen fra en distinkt monofyletisk klad som inkluderer sjimpanse, gorilla og bonobo (ca.7,4 millioner år, 95 % KI: 5,4-9,3 millioner år). Denne topologien er igjen i samsvar med UAS."
 Igjen, under standard fylogeni antas sjimpanse og menneske å være nærmere hverandre enn de er gorilla. Så å finne en seksjon i sjimpanse/bonobo-genomet som ligner mer på gorilla enn på mennesket er et annet eksempel på fylogenetisk inkongruens. Ikke bekymre deg - de bare vifter med tryllestaven UAS for å fikse problemet.
Igjen, under standard fylogeni antas sjimpanse og menneske å være nærmere hverandre enn de er gorilla. Så å finne en seksjon i sjimpanse/bonobo-genomet som ligner mer på gorilla enn på mennesket er et annet eksempel på fylogenetisk inkongruens. Ikke bekymre deg - de bare vifter med tryllestaven UAS for å fikse problemet.
All denne antatte UAS får dem til å foreslå at kromosomfusjonshendelsen skjedde på et tidspunkt da det foregikk artsdannelse helt tilbake i Australopithecus. Det virker ekstremt spekulativt. Hva er UAS, tross alt? Det er en mekanisme som noen ganger påberopes for å bortforklare gensekvenser som ikke passer til standardfylogenien. Her viser rådataene bevis som er fylogenetisk inkongruente. De gjør på en eller annen måte det om til bevis på en fusjonshendelse i Australopithecus.
Bilde 5. Feilaktige forutsetninger -iflg. Davidson
Hva har denne artikkelen vist?
Til slutt ser det ut til at denne artikkelen faktisk viser
*En innviklet historie som involverer en rekke kompliserte mutasjoner er nødvendig for å svare på Fan et al. (2002) sitt spørsmål om hvorfor "fusjonsstedet" ser så "degenerert" ut sammenlignet med hva du ville forvente at et "fusjonssted" skulle se ut som. Det er ikke akkurat en elegant eller overbevisende forklaring, og det får en til å lure på om noe annet foregår her.
 *"Fusjonsstedet" ser ut til å inneholde flere uttrykte funksjonelle gener, inkludert seksjoner som har store effekter på transkripsjon relatert til nevral utvikling. Dette er ikke akkurat hva man ville forvente av et sted som er et resultat av den tilfeldige fusjonshendelsen som deretter opplevde alle slags komplekse mutasjoner!
*"Fusjonsstedet" ser ut til å inneholde flere uttrykte funksjonelle gener, inkludert seksjoner som har store effekter på transkripsjon relatert til nevral utvikling. Dette er ikke akkurat hva man ville forvente av et sted som er et resultat av den tilfeldige fusjonshendelsen som deretter opplevde alle slags komplekse mutasjoner!
Bilde 6. Funksjonell koherens -hva det er
*Fusjonsstedet har forskjellige underseksjoner som er fordelt mellom mennesker og aper i mønstre som motsier standard fylogeni.
Artikkelen påberoper seg forutsigbart UAS for å bortforklare disse fylogenetiske konfliktene - selv om den hevder at dette er bevis på at fusjonshendelsen skjedde under artsdannelse i slekten Australopithecus. Kanskje, men totalt sett er det mye data her som absolutt ikke er hva man ville forvente fra et evolusjonært perspektiv.
 Bilde 7. Casey Luskin
Bilde 7. Casey Luskin
Casey Luskin er geolog og advokat med høyere grad i vitenskap og jus, som gir ham ekspertise i både de vitenskapelige og juridiske dimensjonene av debatten om evolusjon. Han fikk sin doktorgrad i geologi fra University of Johannesburg, og BS- og MS-grader i geovitenskap fra University of California, San Diego, hvor han utstrakt studerte evolusjon, både på hoved- og lavere nivå. Hans jusgrad er fra University of San Diego, hvor han fokuserte studiene på første Amendment, utdanningslov og miljørett.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund